Здравствуй, мой дорогой читатель! Представляю твоему вниманию серию статей про свет для растений. В первой статье разберем подробнее, как происходит поглощение энергии света. Знание об этом процессе позволит на более тонком уровне понимать природу растений.
Итак, приступим. Свет — один из фундаментальных факторов, без которого растение не сможет фотосинтезировать, и, следовательно, развиваться и расти.
Фотосинтез — это процесс образования органических веществ при участии энергии света (происходит восстановление углекислого газа до глюкозы). Органические вещества — это вещества, которые содержат в себе углерод (C) и являются основой всего живого.
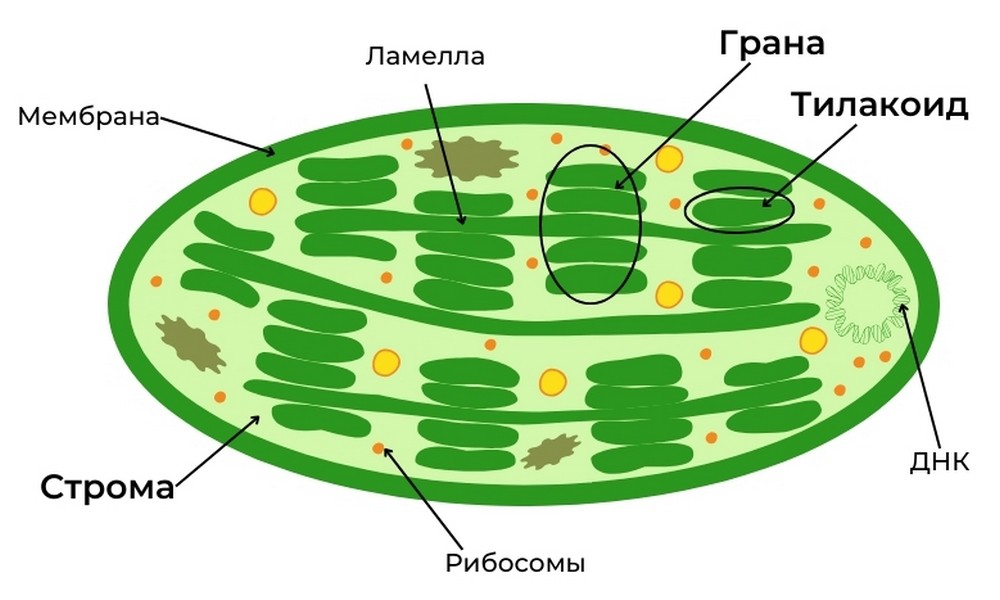
Для этого процесса растительные клетки использует специальные двумембранные органеллы — хлоропласты. В них и протекает весь процесс фотосинтеза.
Фотосинтез делится на светозависимую и светонезависимую фазы, обе из которых важны для растения.
Светозависимая фаза фотосинтеза
Световая фаза протекает в тилакоидах. Тилакоид, в переводе с греческого «мешочек», представляет собой образование внутренней мембраны хлоропластов. Вместе тилакоиды образуют грану.
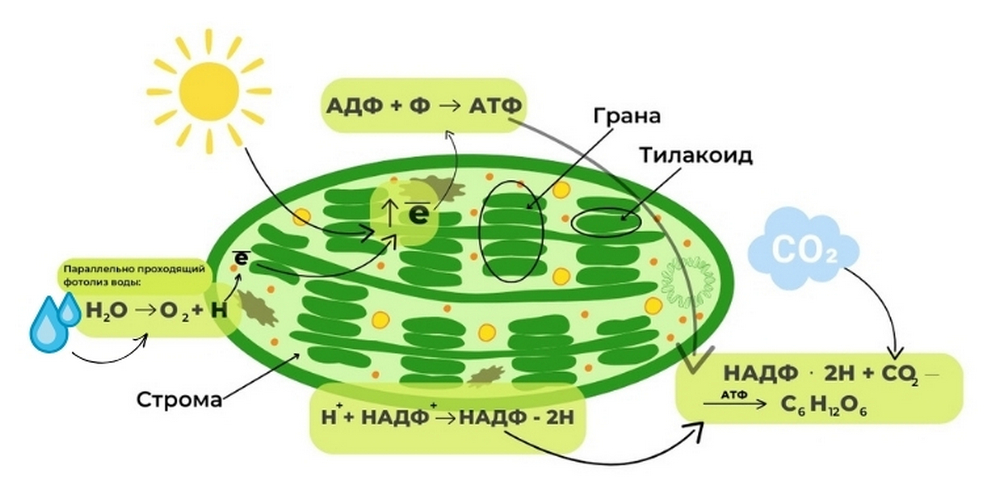
Весь процесс начинается с поглощения солнечного света молекулами хлорофилла. Под воздействием света электроны в этих молекулах переходят в возбужденное состояние (переходят на более высокий энергетический уровень). Это создает энергию, которая необходима для фосфорилирования, т. е. образования АТФ. АТФ — универсальная энергетическая молекула.
Одновременно с этими процессами происходит фотолиз воды — разложение воды под воздействием солнечного света. Вода распадается до кислорода (O2) и водорода (Н). Водород состоит из протона и электрона. Электрон из этого атома соединяется с молекулой хлорофилла, компенсируя тот электрон, который хлорофилл отдал для синтеза АТФ. А протон водорода соединяется с переносчиком НАДФ.
По завершению световой фазы образуются следующие продукты: АТФ, НАДФ∙2Н, а также кислород (О2) — побочный продукт световой фазы фотосинтеза, который улетучивается в окружающую среду.
Светонезависимая фаза фотосинтеза
Темновая фаза протекает в строме — области внутри хлоропласта, окружающей тилакоиды. Используются продукты световой фазы и CO2, получаемый из атмосферы.
Начинается темновая фаза с фиксации углекислого газа. Затем запускается цикл Кальвина, который включается в себя окисление НАДФ∙2H и образование молекул водорода и НАДФ+. Этот цикл представляет собой последовательность химических реакций, в результате которых триозы преобразуются в пентозы, а затем в гексозы, такие как глюкоза.
В процессе цикла Кальвина происходит окисление переносчика НАДФ и высвобождение НАДФ и водорода. Для всех этих реакций требуется энергия АТФ, которая образуется в световой стадии. Благодаря энергии, полученной от АТФ, происходит синтез углеводов и восстановление углекислого газа до глюкозы.
Глюкоза, в свою очередь, является основным источником энергии для многих процессов жизнедеятельности растений. Позднее глюкоза может быть преобразована в целлюлозу или крахмал.
Фотосинтез — это удивительный и сложный процесс, который лежит в основе жизни нашей планеты. Растения, используя световую энергию, воду и углекислый газ, превращают их в органические соединения, обеспечивая питание для себя и других организмов.
Понимание процесса фотосинтеза помогает нам не только лучше понять природу и ее механизмы, но и разработать новые методы улучшения сельского хозяйства, биотехнологии и экологической устойчивости. Благодаря фотосинтезу мы можем наслаждаться красотой природы, а также получать пищу и кислород, необходимые для жизни.
Таким образом, фотосинтез является невероятным процессом, который демонстрирует величие и сложность живой природы, а также важность сохранения экосистем для благополучия нашей планеты и будущих поколений.


